20210730 Ramey Peer Review Chaffee Tool and Plan
To:
Chaffee County Commissioners
P.O. Box 699,
104 Crestone Avenue
Salida, Colorado 81201
From:
Rob Roy Ramey II, Ph.D.
Wildlife Science International, Inc.
P.O. Box 386 Nederland, CO 80466 USA
30 July 2021
Dear Commissioners Baker, Felt, and Granzella,
What follows is my detailed review of the Chaffee County “Wildlife Decision Support Tools for Recreation” (February 20, 2021; hereafter “Tool”) and the “Protect and Restore Wildlife Habitat” portion of the Chaffee County Recreation Management Plan (June 2021; hereafter “Plan”). I direct my comments to you for consideration.
My qualifications:
I earned my Bachelor’s degree in Biology and Natural History from the University of California Santa Cruz, Master’s degree in Wildlife Ecology from Yale University, and Ph.D. in Ecology and Evolutionary Biology from Cornell University. My postdoctoral experience included research at the University of Colorado, Boulder as a USDA postdoctoral fellow, and the Center for Reproduction of Endangered Species at the San Diego Zoo. I have been conducting research on ecology, evolution, and conservation of mountain sheep since 1981. That research includes a wide variety of conservation-related questions on desert, Sierra Nevada, and Rocky Mountain bighorn sheep in North America, and genetics of argali, Urial, and snow sheep in Asia. My doctoral dissertation research was on the population structure, systematics, and evolution of mountain sheep. Later research included host specificity and the evolution of virulence in parasites and bacterial pathogens in bighorn, evolution of the mammalian gut microbiome, as well as the population genetics, phylogenetics, trace element nutrition, population dynamics, and translocation strategies affecting the conservation of North American mountain sheep. With colleagues, I pioneered the live-capture of Argali sheep on the Mongolian steppe, the development of non-invasive fecal DNA technology for genetic research on the endangered Sierra Nevada bighorn sheep, and the concept of metapopulation management to desert bighorn sheep. I have published research on the natural and human-caused factors affecting the population dynamics of bighorn sheep, greater sage grouse, and the critically endangered delta smelt in California
My experience includes work with raptors as well. I surveyed for, observed, and climbed into active nests of peregrine falcons and California condors for research and to recover eggs for captive incubation, at a time when those species were on the brink of extinction. I have climbed to numerous active peregrine falcon nests to aid species recovery, including two high on the face of El Capitan in Yosemite to retrieve eggs thinned by DDE and foster captive reared chicks into the nests. Projects have included collecting eggshells from peregrine nests for DDE research on remote cliffs in Zion and Canyonlands National Parks, and Lake Powell National Recreation Area. At the request of the National Park Service, I made a 12-day ascent to a peregrine nest on the most overhanging section of the northwest face of Half Dome to determine the cause of nest failure. I have also surveyed for and climbed into northwestern goshawk nests to collect prey remains as a wildlife biologist on Inyo National Forest. And for the past 20 years, I have conducted early season observations for Arapaho National Forest to determine the locations of golden eagle nests so that climbing closures may be targeted to just active nests.
As part of my professional duties, I contribute as an ad hoc peer reviewer for a wide variety of scientific journals, as well as habitat conservation plans, endangered species recovery plans, BLM resource management plans, USFS land use plans, environmental impact statements, biological opinions, city and county conservation plans, a habitat exchange habitat quantification tool, research reports, draft scientific manuscripts, grant proposals, and proposed federal rules.
In my spare time, I conduct field research with colleagues on the behavioral ecology, population dynamics, and conservation of African elephants in the northern Namib Desert.
My CV is attached. Ramey CV June 2021
Detailed Review of scientific issues with the Chaffee County Wildlife Decision Support Tools for Recreation (February 20, 2021 version) and the Protect and Restore Wildlife Habitat portion of the Chaffee County Recreation Management Plan (June 2021 version):
The Tool and Plan are based upon a biologically naive and specious assumption.
One of the nine stated objectives of the Plan is to “Stabilize and Enhance Wildlife Populations by 2026.” To achieve this objective both the Tool and the Plan make the fundamental assumption that they must “reduce recreation-related impacts on wildlife and their habitat” because recreation activities in Chaffee County have resulted in unstable populations or population declines. Unfortunately, the stated objective and the underlying assumption on how to achieve it are biologically naive, scientifically unsound, and contrary to the available data and current wildlife management practices. It is apparent to this reviewer, that the authors of the Tool and Plan are unfamiliar with the basics of animal behavior, population biology, and conservation biology.
The fundamental assumption behind the Tool and Plan is both biologically naive and specious because it is assumed that regulating recreation activities will magically result in rebounding wildlife populations in five years. This assumption relies on a simplistic, popular belief that recreational activity is currently the primary limiting factor to wildlife populations in Chaffee County, while ignoring other natural and human-caused factors that decades of research have shown to have a quantitative effect on population dynamics. Briefly, those factors include: disease, predation, competition, invasive species, hunting, regional climatic variation, wildfire, permanent development, and the effects of density-dependence (i.e. population density feedbacks affect population growth rate).
In evaluating the data and scientific literature, the question is not whether animals may react to humans by exhibiting increased vigilance, taking flight, or exhibiting temporarily elevated physiological parameters. The question is whether the animal’s behavioral response(s) ultimately result in a quantifiable effect on their survival and reproduction (Darwinian fitness), and that enough individuals are adversely affected to have a measurable demographic effect on the population, independent of other factors (Wehausen 1980; Gill 2001). Answering that question requires demographic data (population number and trend), plausible cause and effect mechanisms, and the ability to rule out the factors listed above (see Ramey 2012, attached, for an extensive review of the bighorn sheep and human disturbance literature). Fortunately for bighorn sheep and elk in Chaffee County, demographic data exist that can be used to test this hypothesis. And, because these big game populations are hunted, the conditioned reaction of the animals to humans would be expected to be stronger than for unhunted populations (Thorne et al. 1979; Goumas et al. 2020), making it more likely to detect an adverse demographic effect if indeed one exists. For bighorn sheep and elk, these data are examined below.
Bighorn sheep and elk populations and hunter harvest data from Colorado Parks and Wildlife (CPW) annual reports and management plans are contrary to claims made by authors of the Tool and refute its central assumption.
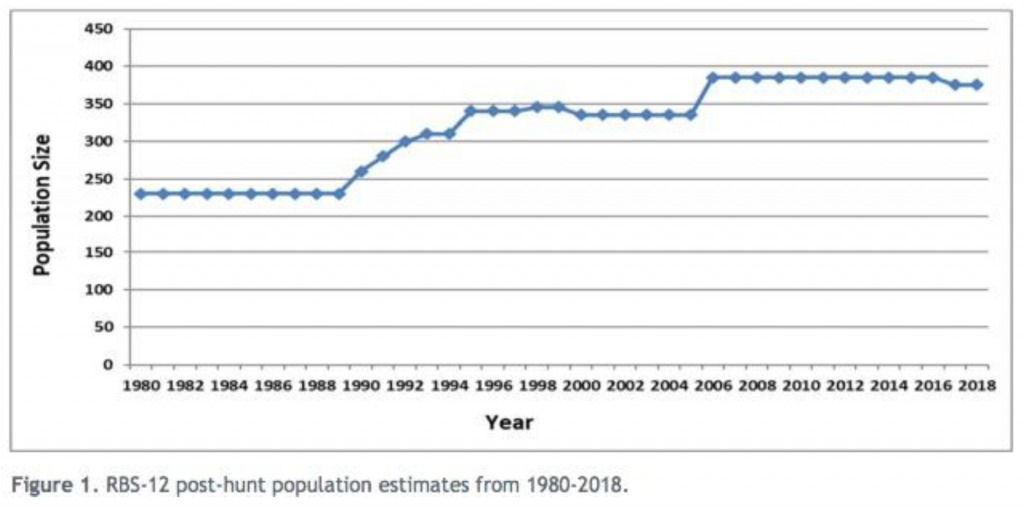
In the case of bighorn sheep for example, CPW’s population data from the Collegiate Range in Data Analysis Unit RBS-12 (GMUs S11, S17, S66, and S76) show that the number of bighorn have consistently increased since 1980 (see Figure 1 from CPW 2020a, below). A post-hunt 2020 population estimate of 375 bighorn is within CPW’s recently-recommended objective of 350-400 individuals (CPW 2020a). It is critical to note that this population increase occurred along with an increase in recreational use and ongoing hunting of the population (with 13-16 bighorn killed out of 25 tags issued annually for the last 3 years; CPW 2020b, 2021a). For bighorn sheep, these data alone clearly refute the central underlying assumption of the Tool and Plan that recreational use in this area has resulted in a bighorn population decline. Below is Figure 1 from CPW (2020a)
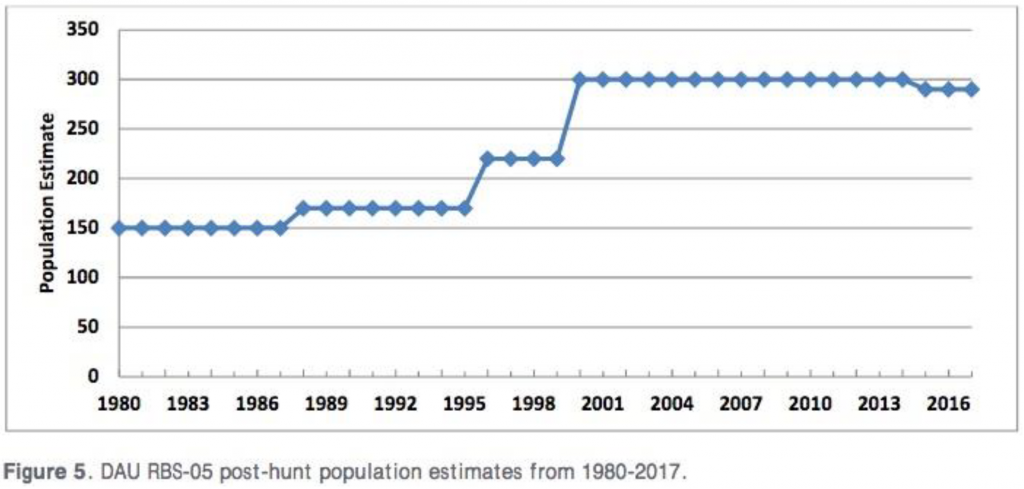
On the East side of the Arkansas Valley, CPW’s population data from the Buffalo Peaks/Mount Silverheels/Tenmile Range bighorn sheep population (Data Analysis Unit RBS-05: GMUs S12, S39, and S78) reveals a similar increase since 1980, from 150 to 300 animals in 2020 (see Figure 5 from CPW 2018, below; and more recent population estimates from CPW 2021a). That increase has occurred despite 15-17 bighorn harvested per year by hunters for the past three years (CPW 2021a).
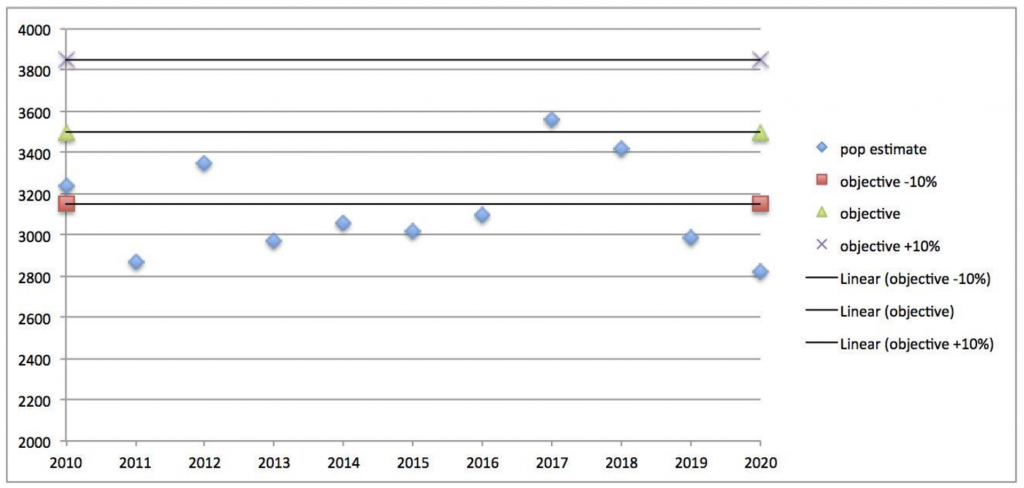
In the case of elk along the Collegiate Range (Data Analysis Unit E-17), this population is intensively managed via hunting by CPW with 400-651 elk killed annually between 2005 and 2015 (or 14-17% of the population, based on CPW’s revised population size estimates), and 334-417 elk estimated to be killed annually by hunters from 2016-2020 (or, 10-12% of the population; CPW 2021b). This does not include the percentage of animals wounded that subsequently die. The 2020 post-hunt elk population was estimated at 2,850 animals, slightly below the lower end of the +/-10% objective of 3,500 elk established in 2011, however, this population has since fluctuated both above and below CPW’s “preferred” population objective since 2010 when estimates of population size were revised with more accurate data and refined population models (data from CPW 2011 and 2021b, see plot below). It is notable that the CPW 2011 population objective was established just as residential development and agricultural fencing to reduce conflicts began to increase. These two factors resulted in the permanent loss of winter range habitat on the Arkansas Valley bottom and consequently reduced the carrying capacity of the elk population (CPW 2011). And, as with hunted elk populations elsewhere, the 2011 CPW management plan for the Collegiate Range reported that, “Elk are often observed seeking refuge in new subdivisions which have created de facto refuges where elk cannot be hunted” (CPW 2011). Important here are the facts that: 1) hunting is itself a form of recreational activity that occurs directly in elk habitat where humans are the predator, with 8,896 to 16,957 elk hunter-days annually from 2005-2020, and 334-615 elk killed annually in E-17 (CPW 2021b); and 2) that hunting has resulted in documented avoidance behavior and modified habitat use in this population (CPW 2011).
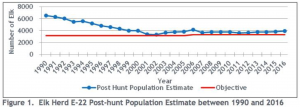
On the east side of the Arkansas Valley, elk in Data Analysis Unit E-22 (GMU 49, 57, 58), is another population intensively managed by CPW. Notably, this elk population has remained stable and fluctuating above CPW’s recommended post-hunt objective of 3,150- 3,500 elk (CPW 2018, see Figure 1 from that report below), despite the increased recreational use in this area (apparently popular with OHVs) and 200-400 animals harvested by hunters each year (CPW 2018, 2021b). In 2020, the post-hunt population estimate was 3,750, again exceeding CPW’s population objective. Therefore, these data refute the central underlying assumption of the Tool and Plan that (non-hunting) recreational use in this area has resulted in an elk population decline.
Finally, mountain goats are not even a native species in Colorado, they are an exotic species that was introduced to Colorado for sport hunting from 1947-1972 when they were erroneously thought to be native and extirpated, and before it was known that they compete with bighorn and harbor pathogens harmful to bighorn (Gross 2001; Lowrey et al. 2018a,b; Mitton 2019; Wolff et al. 2019). For these reasons, the National Park Service (NPS) has been eradicating mountain goats from Rocky Mountain National Park, Olympic National Park, and most recently, from Grand Teton National Park (NPS 2019), consistent with NPS policies on the removal of exotic species that are harmful to native species (NPS 2006). Therefore, it is highly questionable as to why the authors of the Tool included mountain goats as a species of concern instead of eradication when they are an exotic species detrimental to native bighorn sheep. Currently, CPW issues special tags for hunting mountain goats outside of prescribed areas to limit potential colonization of new areas. However, if Chaffee County is serious about bighorn sheep conservation over the long-term, working with CPW to eradicate mountain goats will eliminate competition with, and cross-transmission of infectious disease to, native bighorn sheep.
The CPW data illustrate factually incorrect and misleading statements made by authors of the Tool.
The opening summary paragraph of the Tool makes the bold statement that,
“According to research, 8 of 13 key wildlife populations in Chaffee County — or 65% — are in steady decline. This includes bighorn sheep, down 29% since 2000; mountain goat, down 32% since 2000; and elk, down 11% since 2000. Detailed data provided by Colorado Parks and Wildlife and USFS biologists on these species is available in the Chaffee Recreation Report.”
As an initial matter, comparing this statement to the verifiable sources of CPW data discussed above, the stated declines in bighorn and elk are incorrect. Second, the statement above and paragraph that follows (quoted below), as to cause of purported declines are misleading and provided without any verifiable proof of their claims. More specifically, no “detailed data” can be found in the Chaffee Recreation Report and, as illustrated below, the authors provide no verifiable sources for these and other claims (my emphasis in bold):
“There are multiple factors driving these wildlife population declines, according to wildlife biologists, but there is an increasing body of data from studies across the West that show recreation has measurable negative effects on wildlife, especially in production and winter ranges. Recreation activities displace wildlife, moving them out of high quality to lower quality habitats, these studies show. This reduces the area wildlife use, decreasing the number of animals the landscape can support. Recreation pressure in production areas, from elk calving to raptor nests, has also been shown to decrease the survival of young.”
As noted above for bighorn sheep and elk, there is no data to support the assumption that increasing recreation has resulted in declines of other species in Chaffee County.
Incomplete lists and data missing from the Tool and Plan.
The Plan states, “The wildlife tool is based on information about 44 species…,” however, only 19 species are listed in Table 7 of the Tool (Habitats included in the assessment), along with general categories “fish” and “bats.” Similarly, Table 11 (Recreational effects response function by disturbance level with evidence rating) lists just 15 species as well as “fish,” and two of the species listed on Table 11 are plants, not animals. This begs the immediate questions: What are the other species and information used in developing the Tool and Plan? Where are the data? Neither of these are available on the Envision Chaffee County website nor in the Plan as stated in the Tool. It is worth asking why were the two plants and one insect selected for inclusion, compared to other species?
Biased species and habitat priorities.
The selection of species and habitats included on the list obviously has nothing to do with their actual threat level as species that are “big game” and sport-hunted receive the greatest attention in Tables 7 (habitat types) and 11 (recreational effects response function). Elk are given the highest consideration with 5 habitat types mapped and weighted, mule deer and bighorn with 4 habitat types, and pronghorn with 3 habitat types. Surprisingly, lynx (Lynx canadensis) are erroneously listed as “big game” in Table 7, when in fact this species is federally protected and listed as “threatened” under the Endangered Species Act (unless the authors of the Tool were mistakenly referring to bobcats (Lynx rufus) which are hunted.) These and other mistakes, incomplete listings, lack of citations, missing data, and undocumented methods seriously undermine the scientific credibility of the Tool and Plan. Additionally, the focus on big game that are sport-hunted reveals a willingness by authors of the Tool and Plan, and Planning Commission that certified the Plan, to favor one group of recreational users over others.
Biased data, subjective “corrections,” and murky methods are not the foundations of a robust GIS analysis.
The use of the Great Outdoors Consultants maps of routes and trails appears straightforward. The estimation of recreational use intensity developed for subsequent GIS analyses is murky. This is because it relies on crowd-sourced data from a fitness tracker and geo-location app, STRAVA, typically used by fitness-conscious runners and cyclists. Those data represent a biased sample of users, cited euphemistically by other authors as “where rich white people are” (Eshelman 2020). It is a well-known shortcoming of STRAVA data that it does not capture the recreational use of underserved communities: the economically disadvantaged, aged, and the disabled. As I find no mention of inclusiveness as a goal in the Plan or Tool, it appears that inclusion of these groups is not a priority for Chaffee County officials. Although the Tool mentions modifying the STRAVA data, it appears from the Tool’s description that it was done subjectively by “managers” rather than using an objective, repeatable, and transparent methodology (i.e. Nelson et al. 2021).
Also undocumented was how trail counter data was incorporated (“cross-walked”) with STRAVA data in order to create categorical use variables, especially when these two sources of data were inconsistent with each other, or one was missing. From my experience, trail counters tend to be concentrated near trailheads and consequently tend to over-report trail use farther inland. Simply put, if the data, methods, and code used in analyses are unavailable, then the analysis is not reproducible and therefore it is not science.
An additional question is whether the Great Outdoors Consultants or County’s reliance on private user GIS location data from STRAVA is in violation of Colorado’s recently passed Privacy Protection Act (https://leg.colorado.gov/sites/default/files/documents/2021A/bills/2021a_190_enr.pdf) where individuals have the right to opt-out of the collection of private data (i.e. their locations documented by the fitness app) for which subsequent commercial use is made.
The Animal Habitat Impacts analyses are based upon subjective weightings, buffer distances, disturbance and impact levels, scoring criteria for habitat importance, and recreation effects response values.
It is clear to this reviewer that the Tool’s GIS analysis provides an uncritical reader with a false impression of scientific rigor, when in fact it is lacking.
Arbitrary and capricious thresholds are used in the Tool’s GIS analyses, with no supporting data or scientific research that could be used to compare the thresholds to real-world situations. Starting with the point/line/polygon data layers, these appear to be approximations of potential habitat rather than actual data. Without the data files and associated metadata to pinpoint the source data or opinions used to create the layers it is impossible to verify. Moreover, it has been my experience that polygon layers weight all habitat equally in time and space. In other words, the tendency is to extend polygons to capture all historic locations regardless of how many years ago they were made and how rarely the area is used (see Turner et al. 2004 and 2006). Therefore, such GIS data requires independent validation if it is to be considered “science” and subsequently used for regulatory guidance. In the case of Turner et al. 2004 and 2006, independent reevaluation of bighorn location/GIS data revealed that 66% of critical habitat designated by the U.S. Fish and Wildlife Service was actually non-habitat. That conclusion led to the Court remanding the original critical habitat designation.
Raptor nest buffer distances are not supported by data.
The same argument above holds for raptor buffer distances around points, such as the 800m buffer (1.6km or one mile in diameter) surrounding golden eagle, northern goshawk, peregrine falcon, and prairie falcon nests. This is obviously an oversized, one-size-fits-all buffer that lacks a sound scientific basis (i.e. data that can show a reduced survivorship of individuals or a population-level effect at distances less than this threshold). Therefore, I challenge the authors of the Tool to produce actual data on these species that show nest failure during the nesting season. In fact, none of these species are fragile and the often-repeated myth of human disturbance causing nest abandonment or failure comes from decades in the past (i.e. before the 1970s and the environmental movement). Those early documented cases of “human disturbance” causing nest failure were actually from the destruction of golden eagle nests, killing of young, and shooting of adults from the ground near nests and birds in flight from aircraft. This misguided persecution was carried out by domestic sheep producers and ranchers in the USA (Nelson 1982). In fact, Colorado had a hunting season on golden eagles until 1966. The killing of eagles by Native Americans for feathers used in ceremonial headdresses was another documented form of “human disturbance” (Nelson 1982). During the same period, “human disturbance” of peregrine falcons was from egg collectors who “roped” into nests and mistakenly referred to in the past as “climbers.” And in Scotland and the UK, game keepers shot peregrine falcons on sight to protect game birds (Ratcliffe 1993). I have direct experience of this anti-predator attitude and persecution first-hand, growing up in a rural area in the 1960s and 1970s. Although that dark chapter on persecution of raptors is now closed, some uncritical authors still conflate past human disturbance that had lethal intent with contemporary use of the term “human disturbance” that refers to any human presence in the vicinity of nests, even if is benign.
Experimental data does not support the proposed buffer distances used in the Tool’s GIS analysis.
Experimental evidence reveals a greater tolerance of golden eagles (and other raptors) to human presence and activities than typically parroted in the literature. Three studies on human disturbance of raptors stand out in contrast to the trend described above because they relied on controlled experiments to test the effects of human disturbance on the fitness of raptors (White and Thurow 1985; Holthuijzen et al. 1990; Grubb et al. 2007, 2010). All three utilized disturbances that were clearly threatening (e.g. blasting; threatening approach via foot, vehicle, or helicopter; gunshots and noisemakers), as compared with relatively benign activities such as hiking, rock climbing, and horseback riding. Yet, all three reported a remarkable tolerance of human presence, a decreased response when habituated, and recommended substantially smaller buffer zones than those typically imposed.
Holthuijzen et al. (1990) measured the effects of nearby blasting on nesting prairie falcons, as compared to undisturbed controls. They reported: “This study demonstrated that, in general, blasting had no severe adverse effects on the falcon’s behavioral repertoire, productivity, and occupancy of nesting territories. Therefore, we suggest that when blasting does not occur prior to aerie selection and ceases prior to fledging, blasting that takes place at least 125 m from occupied prairie falcon aeries need not be restricted, provided that peak noise levels do not exceed 140 dB at the aerie (i.e., the noise level we measured for our experimental blasts). We recommend that no more than 3 blasts occur on any given day or 90 blasts during the nesting season.”
White and Thurow (1985) used an experimental approach to quantify the effects of human disturbance on nesting ferruginous hawks. Their “low level” disturbance involved approaching nests on foot while firing a rifle every 20m, driving up to nests, and continuously operating a 3.5hp gasoline motor or noisemaker within 30-50m of a nest. They reported, “Unlike previous reports of substantial nest desertion by raptors as a result of human activity, the number of disturbed nests that were deserted in our study was unexpectedly low.” And, “Our observations suggest that a sufficient buffer zone for brief human disturbance around ferruginous hawk nests is 250 m. Adults will not flush 90% of the time if human activity is confined to distances greater than this.”
Grubb et al. (2007, 2010) directly approached golden eagle nests at close range via helicopter, and quantified behavior and nest success. This study was a poignant refutation to an often repeated but erroneous perception (discussed above) that golden eagles are highly susceptible to human disturbance. The authors reported results contrary to expectations:
“Multiple exposures to helicopters during our experimentation in 2006 and 2007 had no effect on golden eagle nesting success or productivity rates, within the same year, or on rates of renewed nesting activity the following year, when compared to the corresponding figures for the larger population of non- manipulated sites. During our active testing and passive observations, we found no evidence that helicopters bother golden eagles nor disrupt nesting. In 303 helicopter passes near eagles, we observed no significant, detrimental, or disruptive responses. 96% of 227 experimental passes of Apache helicopters at test distances of 0-800 m from nesting golden eagles resulted in no more response than watching the helicopter pass (30%). “
“We found no relationship between helicopter sound levels [even though Apache helicopters were twice as loud as the civilian helicopters] and corresponding eagle ambient behaviors or limited responses, which occurred throughout recorded test levels (76.7-108.8 dB, unweighted).”
“Between all the other aircraft and human activities occurring in the Tri-Canyon Area, as well as their long term coexistance with WPG and apparent indifference to current operations, golden eagles in the area appear acclimated to current levels of activity. “
“For the specific question of WPG operating in the Tri-Canyon Area without potentially impacting nesting golden eagles, we found no evidence that special management restrictions are required. (Authors’ Note: The results of this research were very much unexpected since helicopters are usually considered more disruptive to bald eagles than any other type of aircraft. Plus, golden eagles are traditionally thought to be more sensitive, and therefore more responsive, to human intrusions than bald eagles. However, we found the golden eagles studied during this project to be just as adaptive, tolerant, and acclimated to human activities as any bald eagles in our rather considerable, collective experience with this species. We hypothesize this may at least be in part due to the proximity of the large, growing, and outdoor-oriented population of the Salt Lake Valley and Wasatch Front.”
The experimental data and observations of the authors above are consistent with my extensive experience working with peregrine falcons, golden eagles, and northern goshawks, and entirely inconsistent with the buffer distances used in the Tool.
Subjective weightings, criteria, and response values are not science.
Each one of the weightings, criteria and response values used in the GIS analysis are highly subjective. Those include Disturbance Levels (Table 9), Impact levels (Table 10), Recreational Effects Response Function by Disturbance Level with Evidence Rating, Habitat Rank Components and Scoring Criteria (Tables 12 and 13), and Spatial Criteria for Defining Recreational Disturbance Intensity. All are provided without any citation from scientific literature or presented with a scientifically credible cause and effect mechanism with which to justify them or their scores. Instead, the Tool’s authors attribute these collectively to the input of “Resource Specialists.” Like the lack of listed authors on the Tool and Plan, it is unclear who these “resource specialists” were. However, it is clear is that whoever designed the weightings, criteria, and response values appears to have a weak foundation in animal behavior and population biology. For example, consider the scoring criteria used for Population Trend (Table 12):
“This component captures the local population trend of the associated species: 1 = increasing, 2 = stable or unknown/no data, 3 = slightly declining (<0.5%/yr), 4 = clear decline (0.5-1%/yr), and 5 = strong decline (>1%/yr).”
Note that there are three categories for “local population trend” that involved declines: “slightly declining,” “clear decline,” and “strong decline” with each separated by a half-a-percent change, and any negative change over one percent per year is considered a “strong decline.” There is only one category for “increasing” but with no criteria for identifying if an “increase” has occurred. The author of this does not seem to understand that populations tend to fluctuate to a greater degree annually in both their actual and estimated numbers. Moreover, because it is extremely rare to obtain exact counts, population estimates virtually always have an associated level of uncertainty (expressed in terms of 95% confidence intervals). Therefore, it is extremely unlikely that it would be possible to reliably detect such small changes with any reasonable degree of certainty, and whether such small changes are biologically meaningful at all. By comparison, CPW sets their recommended population objectives within a +/-10% range of an ideal value and do not seem to be concerned outside of that range unless it is a prolonged trend (for examples please refer to the CPW bighorn and elk management plans cited above).
The scientific and biological naiveté of the Tool’s authors becomes increasingly apparent with each new variable introduced in their model. For example, when examining the “disturbance levels” in Table 9, these are criteria based on levels of human disturbance to other humans, not wildlife. The so-called “Evidence” levels (low, moderate, high) in Table 11, leave me with the question: What evidence? There are no citations, no data, and no stated criteria for each level. In short, no evidence. There is no cited scientific research in support of the arbitrary Impact levels and criteria in Table 10, these are clearly subjective. Next, the stated values for Impact by recreation disturbance level in Table 11 are also subjective, as are the Sensitivity & Evidence scoring criteria in Table 12. Subsequently, Habitat Importance scores are compiled in Table 13 as if it is scientifically defensible to simply take five previous arbitrary values and total them to create a new composite value that is even more arbitrary. Values from Tables 11 and 13 for each species are then multiplied and summed to create Composite impact scores used in spatial analyses and create maps to guide decision-makers. If these were observable variables in the real world, with actual data or estimates (with associated confidence intervals), one could account for error propagation. However, that is not possible with the analysis described in the Tool. The GIS analysis above may appear rigorous on the surface, but that veneer of rigor quick disappears once one scratches the surface.
The role of hunting in altering animal behavior is ignored.
It has been my direct experience, conducting research on bighorn sheep across the western USA and Mexico for over 40-years, that bighorn sheep in populations that are hunted react more strongly and at farther distances to human presence than those that are not hunted. More specifically, hunted populations tend to be more vigilant and take flight at greater distances than un-hunted populations, especially during the fall-winter hunting season. The reason is straightforward: in hunted populations, hunters act as predators. Hunters walk directly into the habitat of the bighorn and often for prolonged periods to select and stalk their quarry, then shoot and kill them (with a bow or high-caliber rifle shot), which is likely to be seen by other members of the herd. When humans act as predators on bighorn it conditions the bighorn to view the presence of all humans in their environment as potential predators.
The conditioning of animal behavior through hunting (as well as poaching or culling) has been reported in many other species (Goumas et al. 2020). I have witnessed this in the behavior of North American game species (bighorn, mule deer, elk, pronghorn) as well as in African wildlife from springbok to elephants. Therefore, if a goal of Chaffee County is to have bighorn or elk populations that react less to benign human recreation, the solution is simple: stop hunting them. The same can be said for other species including elk. That is why national parks the world over tend to be populated (and over-populated) with animals that have habituated to sharing their environment with humans exhibiting predictable and non-threatening behaviors. (And for the record, I am not an anti-hunter, I work with sport and trophy hunting organizations in the USA and abroad.)
Similarly, invasive research of bighorn and elk that requires capture via helicopter net- gun or darting with immobilizing drugs and subsequent handling, is functionally equivalent to humans acting as predators. For example, a 2013 bighorn sheep study by CPW required the capturing of pregnant bighorn ewes, resulting in physical restraint, blood drawn, ear tags attached, radio-collars fitted and vaginal implant transmitters inserted in ewes, followed later by the hand-capture, handling, and radio-collaring of their neonatal lambs, just days old. While CPW may assert that bighorn behavior is unaffected, the data suggest otherwise. In the words of the study authors, during that 2013 CPW study, 3 of 15 neonatal lambs captured and radio-collared “died of starvation likely caused by abandonment [by their mothers] after capture” (Grigg et al. 2017).
While such research is important for the management of a species (in that case, respiratory disease transmitted from domestic sheep), from an animal behavior point of view wildlife captures are invasive procedures, similar to predation attempts. Similarly, helicopter pursuit and capture has known deleterious physiological effects, including acidosis and capture myopathy which cause permanent lameness or death, injuries from net-gunning, and behavioral effects such as flight and avoidance of researchers and helicopters (Bleich et al. 1990, 1994). Darting animals with immobilizing drugs may be less intrusive but it is also risky and has resulted in injuries when bighorn tumble down rocky slopes.
Therefore, hunting and research activities cannot reasonably be considered as benign activities without some consequences for subsequent bighorn (or other species) behavioral responses to humans. These activities can create a “landscape of fear” for animals and lead to their increased vigilance and avoidance of humans. In light of these observations, when it comes to human-wildlife interactions that occur with recreation, perhaps the best advice is to educate users of all kinds on how to “not act like a predator.” It has also been proposed that we rethink how we hunt in order to reduce the perception of humans as predators or the association of certain habitats with predation risk (Cromsigt et al. 2013). These are worth further consideration. The opposite approach, to increase the perception of predation risk, can also be used to reduce human wildlife conflicts (i.e. elk in agricultural areas).
There is a body of literature that reports on individual-responses to various recreational activities. However, evidence of a negative population-level effect is notably absent.
To date, the literature on recreationalists disturbance to wildlife (more accurately termed human-wildlife interactions) generally falls into four categories:
- Conjectures about a potential role of human disturbance on population dynamics of the species/population in question but with no actual data showing a demographic effect. Opinions expressed by the authors in the conclusions of their paper are often erroneously cited by subsequent authors, as if these opinions were actual results based on demographic data.
- Measurements of anti-predator behavioral responses (again, with no actual data showing a demographic effect but opinions expressed that they could). The conclusions of these papers generally assume that any observed effect(s) results in a decrease in individual fitness and ultimately population number.
- distances at which animals respond to humans (i.e. exhibit increased vigilance, move away, take flight).
- observations of short-term displacement of animals by humans (using field observations or GPS-collar data);
- measurements of short-term physiological responses to humans (i.e. increased heart rate, cortisol hormone level);
- Correlative studies that compare an increasing “human footprint” (based on GIS data, such as those in the Tool) to trends in habitat utilization or population number. The analytical methods utilized generally fail to rule out other factors that could negatively influence demography. There are papers from other fields of inquiry, such as quantifying effects of energy development (Ramey at el. 2018) or fish hatcheries (i.e. Maunder et al. 2014) on population dynamics, in combination with other factors but not recreational activities.
- Literature reviews that compile reaction distances or recommended buffer zones based upon the opinions of previous authors in the conclusions of their paper. These, in-turn, are melded by authors into their own recommendations, while ignoring methodological issues or weak inferences of the articles cited (i.e. using methods 1,2, and 3 above). The primary issue with this approach is that the final product is just a survey of opinions and subject to confirmation bias.
Reconsidering method #3 above, what if a population remains stable or increases over the long-term along with recreational activity (i.e. 10-20 years with no biologically significant decline)? And, what if this increase continued along with direct sources of mortality such as hunting? Would it then be reasonable to reject the hypothesis that recreational activity is deleterious to the population? I argue that it should be rejected because the data are inconsistent with the hypothesis. This is the current situation in Chaffee County with regards to bighorn sheep and elk. Could this change in the future? It could, given the perfect storm of circumstances. However, at this time there is no sound scientific rationale to use wildlife as the reason to regulate recreation. There may be other reasons to regulate recreational use, type, and intensity, but wildlife species in Chaffee County are not one of those reasons.
For a detailed analysis of the literature on human disturbance of bighorn sheep and how it and bighorn sheep census data was applied to understanding an issue involving recreational trail users in desert bighorn sheep habitat, please refer to testimony provided in Appendix A.
Where is the adaptive management?
The authors of the Tool and Plan make no mention of adaptive management, which is essential to effective management of natural resources. With wolves about to return in number to Colorado through CPW’s reintroduction program, habitat use and population trends of prey species like deer and elk are likely to be affected, making much of the Tool’s habitat mapping obsolete.
Were the contributors to the “Animal Impacts Analysis” of the Tool chosen based on their biological expertise or other criteria?
Returning to the “Disturbance Levels” in Table 9, it is noticeable that the level of noise was included in the evaluation of the “disturbance” to the species examined. While the mating calls of some songbirds has been reported to be obscured by highway noise or oil and gas activity, I am unaware of scientific papers reporting the same effects on the species listed in the Tool. Instead, aesthetic values that are of concern to some user groups appear to have been inserted here.
I note that the Director of the “Quiet Use Coalition” was cited as a contributor to the “Animal Habitat Impacts” section of the Tool. While this individual has many athletic trail running accomplishments, I could find no evidence of qualifications in the fields of animal behavior, ecology, or population biology. He apparently earned his Bachelors degree in biochemistry in 1985 and has had a distinguished trail-running career with numerous awards. According to the Quiet Use Coalition’s website, it “is a spearhead organization of the Quiet Use Movement.” From the Ark Valley Voice (Sep 21, 2019) I found this description,
“As a strong environmental advocacy organization [The Quiet Use Coalition, QUC], says Sobal, it does not merely celebrate or promote nature, it actively works to protect and preserve it. He says QUC will take, and is not afraid to take, strong stances and positions on environmental topics. These are sometimes controversial. QUC has used objections, appeals and lawsuits when necessary to emphasize its positions and obtain needed clarifications on issues.”
The participation of an advocate and litigant as a contributor to the Tool, to the exclusion of other recreational user groups, raises the issue of bias. While recreational activities ranging from hiking 14ers, trail running, cycling, e-biking, horseback riding, motorcycling, OHV riding, and sport hunting may be personally distasteful to some individuals or advocacy groups in Chaffee County, it is clear that these have not been deleterious to the bighorn and elk populations. Therefore, it is worth asking the question, in the absence of data, should wildlife be used as an excuse to push other agendas?
Is Chaffee County on the verge of exceeding its authority?
The authors of the Tool and Plan appear to assume that recreational use on federal lands within Chaffee County can be planned and regulated outside the inclusive public process that is the purview of the National Environmental Policy Act (NEPA). “Input” from a select handful of state and federal staff does not obviate this issue. It is also worth asking why a representative from an organization engaged in advocacy and litigation on local federal decisions was allowed to participate in development of the Tool, without representatives from other stakeholder groups.
I recommend scrapping the current Tool for all of the reasons detailed above. If Chaffee County is serious about contributing to the long-term conservation of its native fish, wildlife, and plant populations, there are other substantive actions that can be taken to address threats to these. A brief list is provided below.
Bighorn sheep
- Work with NGOs and CPW to buy out and retire domestic sheep grazing allotments that overlap or are near native bighorn sheep range. Nearby allotments are identified in CPW (2021a). Transmission of bacterial pathogens from domestic sheep (and goats) to bighorn sheep is a well-documented, long-term threat to bighorn sheep populations (Besser et al. 1994, 2008; Wehausen et al. 2011).
- Reach out to recreational users via news, trailhead information, and social media to report any domestic sheep in or near bighorn sheep range to CPW so they may be quickly removed.
- Consider requiring the reporting of any escaped domestic sheep or goats from hobby farmers or commercial operations to reduce the chance of transmission of bacterial pathogens from domestic sheep and goats to bighorn sheep.
- Incentivize the eradication of mountain goats by CPW. This could be accomplished by providing additional resources for bighorn disease and population monitoring and habitat enhancement, in lieu of lost hunting opportunities for mountain goats. A list of mountain goat populations in and adjacent to Chaffee County may be found in CPW (2021a).
Golden eagles
- Educate and encourage hunters to use non-toxic slugs and shot when hunting, perhaps by subsidizing non-toxic ammunition sales to licensed hunters with tags in Chaffee County. This would reduce the incidence lead contamination/poisoning in golden eagles and other scavengers (Lambertucci et al. 2010).
- Request that road maintenance crews prioritize the removal of carrion from roadways and rights-of-way to reduce the incidence of eagles killed by vehicle strikes.
- Request that electrical service providers identify and mitigate hazardous powerline junctions to reduce electrocution risk to golden eagles.
- Require enhanced wind turbine mortality monitoring and mitigation measures to reduce eagle, raptor, and bat mortalities at these facilities.
Elk and mule deer:
- Work with private land owners to develop and implement wildlife-friendly fencing strategies such as those developed by CPW and funded through the Habitat Partnership Program. Such standards could be built into County land use permitting process and building codes. https://cpw.state.co.us/Documents/LandWater/PrivateLandPrograms/FencingWithWildlif eInMind.pdf
- Work with CPW to estimate different scenarios of projected build-out of private property and how that would affect elk winter range and carrying capacity to inform future land use zoning and permitting.
- To minimize the coming threat of chronic wasting disease (CWD) to mule deer and elk in Chaffee County, instruct appropriate staff to stay current on scientific research and mitigation measures that seek to reduce the spread of CWD prions in the environment. The Prion Research Center at Colorado State University would be a good place to start.
Thank you for this opportunity to comment.
Rob Roy Ramey II, Ph.D.
Literature Cited
Besser TE, Cassirer EF, Potter KA, VanderSchalie J, Fischer A, Knowles DP, Herndon DR, Rurangirwa FR, Weiser GC, Srikumaran S. 2008. Association of Mycoplasma ovipneumoniae infection with population-limiting respiratory disease in free-ranging Rocky Mountain bighorn sheep (Ovis canadensis canadensis). J Clin Microbiol. 46(2):423-30. doi: 10.1128/JCM.01931-07. Epub 2007 Dec 5. PMID: 18057131; PMCID: PMC2238132.
Besser TE, Frances Cassirer E, Highland MA, Wolff P, Justice-Allen A, Mansfield K, Bleich VC, Boyer RT, Pauli AM, Nicholson MC, Anthes RW. 1994. Mountain sheep Ovis canadensis and helicopter surveys: ramifications for the conservation of large mammals. Biological Conservation 70(1):1-8.
Bleich VC, Boyer RT, Pauli AM, Vernoy RL, Anthes RW. 1990. Responses of mountain sheep to helicopter surveys. California Fish and Game 76(4): 197-204.
CPW (Colorado Parks and Wildlife). 2011. Collegiate Range Elk Management Plan Data Analysis Unit E-17 Game Management Units 48, 481, 56, 561. January, 2011. https://cpw.state.co.us/thingstodo/Pages/HerdManagementPlans.aspx
CPW 2019. Bighorn Sheep Herd Management Plan Data Analysis Unit RBS-05, Buffalo Peaks/Mount Silverheels/Tenmile Range, Game Management Units S12, S39, and S78. https://cpw.state.co.us/Documents/Hunting/BigGame/DAU/Elk/E22_DAUPlan_June201 8.pdf
CPW 2018. Buffalo Peaks Elk Management Plan Extension Data Analysis Unit E-22 Game Management Units 49, 57, 58. June 2018. https://cpw.state.co.us/Documents/Hunting/BigGame/DAU/Elk/E22_DAUPlan_June201 8.pdf
CPW 2020a. Collegiate Range Bighorn Sheep Herd Management Plan, Data Analysis Unit RBS-12, Game Management Units S11, S17, S66, and S76. https://cpw.state.co.us/Documents/Hunting/BigGame/DAU/BighornSheep/RBS12DAUPl an_CollegiateRange.pdf
CPW 2020b. Colorado Rocky Mountain Bighorn Sheep 2020 Posthunt Population Estimates – Draft 12/09/2020. https://cpw.state.co.us/Documents/Hunting/BigGame/Statistics/RockyMountainBighornS heep/2020RockyMtnBighornPopulationEstimates.pdf
CPW 2021a. Rocky Mountain Bighorn Sheep Hunting Statistics. https://cpw.state.co.us/thingstodo/Pages/Statistics-Rocky-Mountain-Bighorn-Sheep.aspx
CPW 2021b. Elk Hunting Statistics. https://cpw.state.co.us/thingstodo/Pages/Statistics-Elk.aspx
Cromsigt JPGM, Kuijper DPJ, Adam M, Beschta RL, Churski M, Eycott A, Kerley GIH, Mysterud A, Schmidt K, West K. 2013. Hunting for fear: innovating management of human–wildlife conflicts. Journal of Applied Ecology 50(3):544-549. https://doi.org/10.1111/1365-2664.12076
Davis MA, Foreyt W. 2013. Bighorn sheep pneumonia: sorting out the cause of a polymicrobial disease. Prev Vet Med. 2013 Feb 1;108(2-3):85-93. doi: 10.1016/j.prevetmed.2012.11.018. Epub 2012 Dec 17. Erratum in: Prev Vet Med. 2013 May 1;109(3-4):185. PMID: 23253148.
Eshelman E. 2020. Can You Hear Me Now? The Vulnerability of Cellular and Smartphone Use on the Battlefield. Master of Science Thesis in Information Strategy and Political Warfare. The Naval Postgraduate School, Monterey, CA. Available: https://apps.dtic.mil/sti/pdfs/AD1114556.pdf
Gill JA, Norris K, Sutherland WJ. 2001. Why behavioural responses may not reflect the population consequences of human disturbance. Biological Conservation 97:265-68.
Goumas M, Lee VE, Boogert NJ, Kelley LA, Thornton A. 2020. The role of animal cognition in human-wildlife interactions. Frontiers in Psychology 11:589978. doi: 10.3389/fpsyg.2020.589978.
Gross JE. 2001. Evaluating effects of an expanding mountain goat population on native bighorn sheep: a simulation model of competition and disease. Biological Conservation 101(2):171-185. ISSN 0006-3207, https://doi.org/10.1016/S0006-3207(01)00062-3
Grubb TG, Delaney DK, Bowerman WW. 2007. Investigating Potential Effects of Heli- Skiing on Golden Eagles in the Wasatch Mountains, Utah. Final Report to the Wasatch- Cache National Forest, Study No. RMRS-RWU-4251-P2-2, Agreement No. 05-JV- 11221607-237, USDA Forest Service, Rocky Mountain Research Station.
Grubb TG, Delaney DK, Bowerman WW, Wierda MR. 2010. Golden eagle indifference to heli-skiing and military helicopters in northern Utah. Journal of Wildlife Management 74(6):1275-1285.
Holthuijzen AMA, Eastland WG, Ansell AR, Kochert MN, Williams RD, Young LS. 1990. Effects of Blasting on Behavior and Productivity of Nesting Prairie Falcons. Wildlife Society Bulletin 18(3):270-281
Lambertucci SA, Donázar JA, Hiraldo F. 2010. Poisoning people and wildlife with lead ammunition: Time to Stop Environmental Science and Technology 44: 7759–7760.
Lowrey B, Butler CJ, Edwards WH, Wood ME, Dewey SR, Fralick GL, Jennings-Gaines J, Killion H, McWhirter DE, Miyasaki HM, Stewart ST, White KS, White PJ, Garrott RA. 2018a. A Survey of Bacterial Respiratory Pathogens in Native and Introduced Mountain Goats (Oreamnos americanus). Journal of Wildlife Diseases 54(4):852-858. doi: 10.7589/2018-02-025. Epub 2018 Jun 14. PMID: 29902131.
Lowrey B, Garrott RA, McWhirter DE, White PJ, DeCesare NJ, Stewart ST. 2018b. Niche similarities among introduced and native mountain ungulates. Ecological Applications (5):1131-1142. doi: 10.1002/eap.1719. Epub 2018 Apr 26. PMID: 29573503.
Mitton J. 2019. Introduced mountain goats have colonized much of the land above the trees. Colorado Arts and Sciences Magazine. University of Colorado Boulder. https://www.colorado.edu/asmagazine/2019/12/06/introduced-mountain-goats-have- colonized-much-land-above-trees.
Nelson T, Ferster C, Laberee K, Fuller D, Winters M. 2021. Crowd-sourced data for bicycling research and practice, Transport Reviews 41:1, 97-114, DOI: 10.1080/01441647.2020.1806943
NPS (National Park Service). 2006. Management Policies (Section 4.4.4.2). U.S. Department of the Interior, National Park Service. https://www.nps.gov/policy/MP_2006.pdf
NPS (National Park Service). 2019. Finding Of No Significant Impact Mountain Goat Management Plan Environmental Assessment. https://bloximages.newyork1.vip.townnews.com/jhnewsandguide.com/content/tncms/ass ets/v3/editorial/3/77/3776b336-5f26-5bf5-ae8e-8f0bdd4b1ad4/5dafc14a431ef.pdf.pdf
Raghavan B, Erickson K, Kugadas A, Batra SA, Call DR, Davis MA, Foreyt WJ, Srikumaran S. 2016. Role of carriers in the transmission of pneumonia in bighorn sheep (Ovis canadensis). Biol Open. 2016 Jun 15;5(6):745-55. doi: 10.1242/bio.018234. PMID: 27185269; PMCID: PMC4920194.
Ramey RR. 2012. Testimony on Senate Bill AB880. Before the State Senate Committee on Natural Resources and Wildlife, California State Legislature. July 3, 2012. 10 pages.
Thorne, ET, Varcalli T, Becker K, Butler GB. 1978. Some Thoughts on the Consequences of Non-trophy Sheep Hunting in the Wind River Mountains of Wyoming. Proceedings of the Northern Wild Sheep and Goat Conference. http://www.nwsgc.org/contents/1978contents.html
Turner JC, Douglas CL, Hallum CR, Krausman PR, Ramey RR. 2004. Determination of critical habitat for the endangered Nelson’s bighorn sheep in southern California. Wildlife Society Bulletin 32(2):427-448.
Turner JC, Douglas CL, Hallum CR, Krausman PR, Ramey RR. 2006. Ostermann’s assumption of a flawed habitat model is premised on facts not in evidence: Turner et al. (2005) response to Ostermann et al. (2005). Wildlife Society Bulletin 33(4):1465–1473.
Wehausen JD.1980. Sierra Nevada bighorn sheep: history and population ecology. Doctoral dissertation. University of Michigan, Ann Arbor.
Wehausen JD, Kelley ST, Ramey RR. 2011. A review of experimental evidence concerning respiratory disease transmission between domestic sheep and bighorn sheep. California Fish and Game 97(1):7-24.
White CM, Thurow TL. 1985. Reproduction of ferruginous hawks exposed to controlled disturbance. The Condor 87:14-22.
Wolff PL, Blanchong JA, Nelson DD, Plummer PJ, McAdoo C, Cox M, Besser TE, Muñoz-Gutiérrez J, Anderson CA. 2019. Detection of Mycoplasma ovipneumoniae in Pneumonic Mountain Goat (Oreamnos americanus) Kids. Journal of Wildlife Diseases 55(1):206-212. doi: 10.7589/2018-02-052. Epub 2018 Aug 30. PMID: 30161017
Wood ME, Fox KA, Jennings-Gaines J, Killion HJ, Amundson S, Miller MW, Edwards WH. 2017. How Respiratory Pathogens Contribute to Lamb Mortality in a Poorly Performing Bighorn Sheep (Ovis canadensis) Herd. Journal of Wildlife Disease 53(1):126-130. doi: 10.7589/2016-05-097. Epub 2016 Sep 30. PMID: 27690193.
Appendix A.
Testimony on AB880 (A bill to reopen the Bump and Grind Trail, subsequently approved by the legislature with bi-partisan support and signed by Governor Jerry Brown.)
Before the State Senate Committee on Natural Resources and Wildlife, California State Legislature
Prepared by
Rob Roy Ramey II, Ph.D.
(Member, IUCN Caprinae Specialist Group) Wildlife Science International, Inc.
P.O. Box 386 Nederland, CO 80466
3 July 2012
Honorable Senators. Good Morning. I’m here today to provide some accurate information on desert bighorn sheep (Ovis canadensis nelsoni). These animals are not fragile creatures or susceptible to human disturbance as they have been portrayed to be. Instead, these animals live in extreme environments, are well adapted to avoid predation by mountain lions, coyotes, bobcats, and golden eagles. And where human activities are predictable and non-threatening, bighorn sheep readily habituate to human activity. They are already habituated to human activity in the Coachella Valley; they have commonly and repeatedly entered busy human environments such as major golf resorts, popular hotels, and the most widely used traffic arteries in the Coachella Valley.
In this field, there is a complete absence of a clearly defined, scientifically defensible, causal link between human disturbance and reduced bighorn survival or habitat abandonment resulting in population decline. The main sources of decline of bighorn sheep populations are factors such as predation, rainfall, and disease, all of which are independent of the number of hikers. In fact, the only experimental research that actually tested, instead of speculating on, a population response noted that the population increased as the number of hikers increased. The hypothesis that human disturbance has had demographic effect on bighorn sheep populations lacks factual support. The same can be said of the unpublished Joshua Tree National Monument study (Thompson et al. 2007). Bighorn sheep made adjustments in their use patterns in response to increased human activity on weekends and readjusted their use patterns after the people had left. No demographic effect of human disturbance was found, like other studies on the subject of human disturbance the authors merely speculated that it could potentially occur.
In contrast, hundreds of thousands of visitors pass close to the desert bighorn sheep exhibit at the Living Desert Reserve and the ewes must be give birth control to keep them from reproducing.
The State’s own data shows that the bighorn sheep population is steadily increasing in the North Santa Rosa Mountains area, despite the increasing use of this particular trail (The Bump and Grind Trail).
The recent CDFG population estimates for the North Santa Rosa Mountains: 2006 – 49 bighorn*
2008 – 77 bighorn*
2010 – 90 bighorn **
*February 25, 2009 memo on PBS population estimates from CDFG biologists Randy Botta and Steve Torres, CDFG.
** January 14, 2011 Results of the 2010 bighorn sheep helicopter survey in the Peninsular Ranges of southern California (Randy Botta, CDFG).
The State permits the hunting of the bighorn sheep in the northern Coachella Valley while they’re considered endangered in the Southern part; and it is the same subspecies (Wehausen and Ramey 1993). State governments in Arizona, New Mexico, Nevada, and Utah permit these sheep to be hunted. The California State Government permits ewes, including ewes with lambs, to be chased down with helicopter and net-gunned at close range, subdued and hobbled, fitted with radio collars, or slung below helicopters or loaded into horse trailers prior to their being taken to a new area for release. The State Government has allowed the hand-capture of small lambs in lambing areas in order for them to be fitted with radio collars. This is considered to be an acceptable risk, while trail use by hikers is considered to be an unacceptable risk.
Human disturbance
I have reviewed the entire published literature on the subject of human disturbance and bighorn sheep, and it is almost entirely based on opinion without supporting experimental evidence or rigorous hypothesis testing (see literature cited). For instance, 26 of the papers relied on unsubstantiated opinion or interpretation of limited or anecdotal observations to support conclusions regarding human disturbance. None demonstrated decreased fitness of individuals or populations as a result of human disturbance. Similarly, none documented any permanent abandonment of range due to transient human disturbance, and any apparent displacement was temporary (Blong and Pollard 1968; Jorgenson 1974; Deforge 1972; MacArthur et al. 1979; Graham 1980; Leslie and Douglas 1980; Wilson et al. 1980; Campbell and Remington 1981; Purdy and Shaw 1981; Cunningham 1982; Deforge 1982; DeForge et al. 1982; MacArthur et al. 1982; Holt and Bleich 1983; Wehausen 1983; Cunningham and Omart 1986; DeForge et al. 1995, 1997; Etchberger et al. 1989; Boyce at al. 1992; Harris 1992; McCarthy and Bailey 1994; Rubin et al. 1998; Etchberger and Krausman 1999; Wagner 1999; Rubin et al. 2000). Seven papers that measured flight or movement response to humans were only able to suggest a limited and transitory behavioral response to human activity over short distances. Again, none documented any permanent abandonment of range due to transient human disturbance, and any apparent displacement was temporary and of no demographic consequence (Hicks and Elder 1979; Hamilton et al. 1982; King and Workman 1986; Papouchis et al. 1999; Rubin et al. 2002; Keller and Bender 2007; Thompson et al. 2007).
The only paper that utilized an experimental design to measure demographic effects of human activity on bighorn sheep reported that the bighorn sheep population actually increased along with the number of hikers in a particular area (Wehausen 1980). Demographic data show that the same increase in bighorn sheep along with the number of hikers in an area is also occurring in the North Santa Rosa Mountains. A bighorn population increase is also occurring in the southern part of the Carrizo Canyon subpopulation where human use has increased over the last decade from illegal immigrants, drug smugglers, and Border Patrol Agents in the vicinity of Interstate 8. Similarly, experience with construction and mining projects in or near bighorn sheep habitat (arguable several orders of magnitude larger and more permanent than a hiking trail), including highway construction and maintenance (e.g. Hwy 74), transmission lines (e.g. Palo Verde-Devers No. 1 transmission line, in Arizona (Smith et al. 1986)), and mining (Wehausen 1980; Andaloro and Ramey 1981; Oehler et al. 2005) have not been shown to result in bighorn sheep population declines. And contrary to expectations, Oehler et al. 2005 reported that mountain lion predation was lower near the active mine than in the undisturbed area away from it. Therefore, much of what has passed for the scientific on the human disturbance of bighorn sheep, has been nothing more than unsupported opinion and speculation on what the effects might be. Like competition, human disturbance is only of importance if it has a negative demographic effect on populations, and such an effect has not been found.
Based on an understanding of plausible cause and effect mechanisms, so long as a few reasonable precautions are taken, effects of trail hiking on bighorn sheep will be minimal or non-existent. For example, the presumed level of risk of ewes abandoning lambs is frequently overstated. Even during the lambing season, there is little risk of ewes permanently abandoning lambs. This is because ewes contribute a substantial parental investment in gestation and rearing and, as a result, the probability of ewes abandoning lambs under any circumstance is extremely remote. In fact, researchers at the Bighorn Institute in Palm Springs California, regularly caught young lambs by hand or with hand- held nets to attach radio collars to them and reported no problems with abandonment (J. DeForge, personal communication). However, if they are flushed from steep escape terrain in a lambing area, very young lambs (less than 2-3 weeks old) can be placed at risk to predation and injury from falls. As a result, measures to limit access directly into an active lambing area may be justified but there is no credible scientific justification at all for limiting access to the viewshed of the surrounding area, or to lamb-rearing habitat.
Access to water during the hot, dry summer months, in low elevation mountain ranges is necessary for desert bighorn sheep survival. Therefore, seasonal restrictions or rerouting of trails in the immediate vicinity of water may be reasonable where water sources are few or limited. However, it is important to realize that bighorn sheep may use alternative sources of water or adjust the time they access water. In areas where bighorn sheep are habituated to humans such restrictions may not be necessary. Such restrictions are unnecessary on the Bump and Grind trail as there is no water in the vicinity.
Lambing area vs. lamb rearing areas
I now to address the issue of what constitutes a lambing area vs. lamb rearing area, as the two are quite different. Lambing areas are steep and rugged patches of habitat where ewes go to give birth and raise their lambs in relative safety during their first few weeks of life. More precisely, lambing habitat is defined as an adequacy of expanse (>2 ha) to provide escape from predation for the pre-parous female and the postpartum female and neonate within 1 km of perennial water (Smith et al. 1991; Johnson and Swift 2000; Singer et al. 2000a,b; Zeigenfuss et al. 2000; Turner et al. 2004). They typically use these areas for 2-3 weeks until the lambs are agile and the risk of predation is consequently lower. At that time, ewes and lambs expand their range into lamb-rearing habitat which is essentially ewe summer range. In lamb rearing habitat ewes frequently form groups with other ewes and their lambs, which provides greater vigilance and increased safety in numbers. However, lactating ewes are still constrained by the need for water, especially during the hot summer months. Lambs are subsequently weaned at 3 and 5 months depending upon forage quality. Lamb survival is highly variable and driven by precipitation, predation, and disease, all of which are independent of levels of human activity, such as trail hiking.
While restrictions on access directly into lambing areas are justified, there is no published literature that suggests that lamb-rearing habitat requires similar restrictions, except when water sources are very limited and only in the immediate vicinity (see expanded discussion below). Bump and Grind Trail is not a lambing area, nor is it a favorable lamb-rearing habitat, as evidenced by the fact that only a handful of sightings exist in the area over the past 40 years. And just because a ewe and a lamb (e.g., > one month old) were recently observed crossing a trail, does not make it a lambing area.
Bighorn sheep often pass through while exploring their environment or moving between one favorable habitat patch and another. Lambing areas are not created by administrative fiat, they are defined by data: steep slopes >80%, rugged terrain, and the documented presence of multiple ewes with small lambs (<2-3 weeks old). Such data are lacking for the Bump and Grind Trail. The nearest lambing area is across a valley and above Magnesia Spring, the greatest concentration of lambing is in Bradley canyon. A suspect lambing area south of Ramon Peak is not yet confirmed by data.
While it could be argued that lambing season extends from January through June, in fact few lambs are born at either end of that period. The data show that 78 to 100% of lambs in the Peninsular Ranges are actually born during February–April each year (Rubin et al. 2000). Lambs that are born early or late in the season have lower survival rates than those born peak season (due to Natural Selection).
The trail opponents are well intentioned and share our collective concern for the well being of the desert bighorn sheep, but are simply mistaken on the science on this issue, and we should not allow antiquated assumptions and beliefs to pass as scientific understanding. The bottom line is that there is no scientific support–at all–for the closure of this trail.
Thank you for this opportunity to comment.
Literature Cited
Bleich, V.C., R.T. Boyer, A.M. Pauli, M.C. Nicholson, and R.W. Anthes. 1994. Mountain sheep Ovis canadensis and helicopter surveys: ramifications for the conservation of large mammals. Biological Conservation 70(1):1-8.
Bleich, V.C., R.T. Boyer, A.M. Pauli, R.L. Vernoy, and R.W. Anthes. 1990. Responses of mountain sheep to helicopter surveys. California Fish and Game 76(4): 197-204.
Blong, B. and W. Pollard. 1968. Summer water requirements of desert bighorn in the Santa Rosa Mountains, California, in 1965. California Fish and Game 54(4):289-296.
Boyce, W.M., T. Bunch, S. Cunningham, J. DeForge, D. Jessup, and J. Wehausen. 1992. Stress and bighorn sheep. Panel Discussion. Desert Bighorn Council Transactions.
Campbell, B.H., and R. Remington. 1981. Influence of construction activities on water- use patterns of desert bighorn sheep. Wildlife Society Bulletin 9:63-65.
Cunningham, S.C. 1982. Aspects of the ecology of Peninsular desert bighorn sheep (Ovis canadensis cremnobates) in Carrizo Canyon, California. Unpublished Masters Thesis. Arizona State University.
Cunningham, S.C. and R.D. Ohmart. 1986. Aspects of the ecology of desert bighorn sheep in Carrizo Canyon, California. Desert Bighorn Council Transactions 14-19.
Dunaway, D. J. 1971. Human disturbance as a limiting factor of Sierra Nevada bighorn sheep. Transactions of the North American Wild Sheep Conference 1:165-173.
DeForge, J.R. 1972. Man’s invasion into the bighorn’s habitat. Desert Bighorn Council Transactions 20:15-17.
DeForge, J.R. 1980. Population biology of desert bighorn sheep in the San Gabriel Mountains of California. Desert Bighorn Council Transactions 24:29-32.
Deforge, J.R., S.D. Osterman, C.W. Willmott, K.B. Brennen, and S.G. Torres. 1997. The ecology of Peninsular bighorn sheep in the San Jacinto Mountains, California. Desert Bighorn Council Transactions 41:8-25.
Duncan, G. E. 1960. Human encroachment on bighorn habitat. Desert Bighorn Council Transactions 4:35-37.
Etchberger, R.C., Krausman, P.R., and R. Mazaika. 1989. Mountain sheep habitat characteristics in the Pusch Ridge Wilderness, Arizona. Journal of Wildlife Management 53(4):902-907.
Etchberger and P.R. Krausman. 1999. Frequency of birth and lambing sites of a small population of mountain sheep. Southwestern Naturalist 44(3):354-360.
Ferrier, G. J. 1974. Bighorn sheep along the lower Colorado River: 1974-2050. Desert Bighorn Council Transactions 18:40-45.
Graham, H. 1980. The impact of modern man. Pages 288-309 in: The desert bighorn: its life history, ecology, and management. G. Monson and L. Sumner, eds. University of Arizona Press, Tucson, AZ.
Hamilton, K. S., S. A. Holl, and C. L. Douglas. 1982. An evaluation of the effects of recreational activity on bighorn sheep in the San Gabriel Mountains, California. Desert Bighorn Council Transactions 26:50-55.
Harris, Lisa Kim. 1992. Recreation in mountain sheep habitat. Doctoral dissertation, University of Arizona.
Hicks, L. L., and J. M. Elder. 1979. Human disturbance of Sierra Nevada bighorn sheep. Journal of Wildlife Management 43:909-915.
Holl, S.A. and V.C. Bleich. 1983. San Gabriel Mountain sheep: biological and management considerations. USDA, San Bernadino National Forest, San Bernadino, CA.
Johnson, T.L., and D.M. Swift. 2000. A test of a habitat evaluation procedure for Rocky Mountain bighorn sheep. Restoration Ecology 8:47–56.
Jorgensen, P. 1974. Vehicle use at a desert bighorn watering area. Desert Bighorn Council Transactions 18:18-24.
Keller, B.J. and L.C. Bender. 2007. Bighorn Sheep Response to Road-Related Disturbances in Rocky Mountain National Park, Colorado. Journal of Wildlife Management 71(7):2329-2337.
King, M.M., and G.W. Workman. 1982. Desert bighorn on BLM lands in southeastern Utah. Desert Bighorn Council Transactions 26:104-106.
Krausman, P.R. and J.J. Hervert. 1983. Mountain sheep responses to aerial surveys. Wildlife Society Bulletin 11(4):372-375.
Kovach, S. D. 1979. An ecological survey of the White Mountain Peak bighorn. Desert Bighorn Council Transactions 23:57-61.
Krausman, P.R., W.W. Shaw, and J.L. Stair. 1979. Bighorn sheep in the Pusch Ridge Wilderness Area, Arizona. Desert Bighorn Council Transactions 23:40-46.
Krausman, P.R., and R.C. Etchberger. 1995. Response of desert ungulates to a water project in Arizona. Journal of Wildlife Management 59:292-300.
Leslie, K.M., Jr. 1977. Home range, group size, and group integrity of the desert bighorn sheep in the River Mountains, Nevada. Desert Bighorn Council Transactions 21:25-28.
Leslie, D.M., Jr., and C.L. Douglas. 1980. Human disturbance at water sources of desert bighorn sheep. Wildlife Society Bulletin 8:284-442.
MacArthur, R.A., Johnston, R.H., and V. Geist. 1979. Factors influencing heart rate in free-ranging bighorn sheep: a physiological approach to the study of wildlife harassment. Canadian Journal of Zoology 57:2010-2021.
MacArthur, R.A., Geist, V., and R.H. Johnston. 1982. Journal of Wildlife Management 46(2):351-358.
MacArthur, R.A., Geist, V., and R.H. Johnston. 1986. Cardiac responses of bighorn sheep to trapping and radio instrumentation. Canadian Journal of Zoology 64(5):1197- 1200.
Martucci, R.W., D.A. Jessup, G.A. Gronert, J.A. Reltan, and W.E. Clark. 1992. Blood gas and catecholamine levels in capture stressed desert bighorn sheep. Journal of Wildlife Diseases 28(2):250-254.
McCarthy, C.W. and J.A. Bailey. 1994. Habitat requirements of desert bighorn sheep. Colorado Division of Wildlife Terrestrial Wildlife Research Special Report Number 69.
Nelson, M. 1966. Problems of recreational use of game ranges. Desert Bighorn Council Transactions 10:13-20.
Nelson MW. 1982. Human impacts on golden eagles: a positive outlook for the 1980s and 1990s. Raptor Research 16(4):97-103.
Papouchis, C.M., F.J. Singer, and W.B. Sloan. 1999. Effects of increased recreational activity on desert bighorn sheep in Canyonlands National Park, Utah. Pages 364-391 In Singer, F. J., and M. A. Gudorf. (eds). Restoration of bighorn sheep metapopulations in and near 15 national parks: Conservation of severely fragmented species. Volume III: Research Findings. U.S. Geological Survey Open File Report 99-102, Midcontinent Ecological Science Center, Fort Collins, Colorado. 391pp.
Purdy, K.G., and W.W. Shaw. 1981. An analysis of recreational use patterns in desert bighorn habitat: The Pusch Ridge Wilderness case. Desert Bighorn Council Transactions 25:1-5.
Ratcliffe, D. A. 1993. The peregrine falcon. Second edition. T & AD Poyser, London, England.
Rubin, E.S., W.M. Boyce, M.C. Jorgensen, S.G. Torres, C.L. Hayes, C.S. O’Brien, and
D.A. Jessup. 1998. Distribution and abundance of bighorn sheep in the Peninsular Ranges, California. Wildlife Society Bulletin 26(3):539-551.
Rubin, E.S., W.M. Boyce, and V.C. Bleich. 2000. Reproductive strategies of desert bighorn sheep. Journal of Mammalogy 81(3):769-786.
Rubin, E.S, W.M. Boyce, C.J. Stermer, and S.G. Torres. 2002. Bighorn sheep habitat use and selection near and urban environment. Biological Conservation 104:251-263.
Singer, F.J., V.C.Bleich, and M.A. Gudorf. 2000a. Restoration ofbighorn sheep metapopulations in and near western national parks. Restoration Ecology 8:14–24.
Singer, F.J., C.M. Papouchis, and K.K. Symonds. 2000b. Translocations as a tool for restoring populations of bighorn sheep. Restoration Ecology 8:6–13
Smith, T. S., J.T. Flinders, and D. S.Winn. 1991. A habitat evaluation procedure for Rocky Mountain bighorn sheep in the intermountain west. Great Basin Naturalist 51:205–225.
Turner, J.C., C.L. Douglas, C.R. Hallum, P.R. Krausman, and R.R. Ramey (2004) Determination of critical habitat for the endangered Nelson’s bighorn sheep in southern California. Wildlife Society Bulletin. 32(2):427-448.
Thompson, D., K. Longshore, and C. Lowrey. 2007. The impact of human disturbance on desert bighorn sheep (Ovis canadensis nelsoni) in the Wonderland of Rocks / Queen Mountain region of Joshua Tree National Park, California. Unpublished final report prepared for Joshua Tree National Park, CA. 16 May 2007. University of Nevada, Las Vegas.
Van Den Akker, J.B. 1960. Human encroachment on bighorn habitat. Desert Bighorn Council Transactions 4:38-40.
Wagner, G. 1999. Activity patterns of Rocky Mountain bighorn ewes in central Idaho. In. Second North America Wild Sheep Conference. April 6-9 1999. Reno, NV. 103-121.
Wehausen, J.D. 1979. Sierra Nevada bighorn sheep: an analysis of management alternatives. Cooperative admin. Report. Inyo National Forest and Sequoia, Kings Canyon, and Yosemite National Parks. 92pp.
Wehausen, J.D. 1980. Sierra Nevada bighorn sheep: history and population ecology. Doctoral dissertation, University of Michigan, An Arbor.
Wehausen, J.D. 1983. White Mountain bighorn sheep: an analysis of current knowledge and managemement alternatives. Administrative Report, Inyo National Forest, contract 53-9JC9-0-32.
Wilson, L.O., J. Blaisdell, G. Welsh, R. Weaver, R. Brigham, W. Kelly, J. Yoakum, M. Hinks, J. Turner, and J. DeForge. 1980. Desert bighorn habitat requirements and management recommendations. Desert Bighorn Council Transactions 1-7.
Zeigenfuss, L.C., F. J. Singer, and M.A.Gudorf. 2000. Test of a modified habitat suitability model for bighorn sheep.